Funded by the U.S. National Science Foundation
As major consumers and remineralizers of bacterial and phytoplankton biomass, phagotrophic protists play a central role in the structure and functioning of marine food webs. Protistan grazers can show complex feeding responses, including selective ingestion or avoidance, based on characteristics of prey cells. Such selectivity has the potential to affect carbon cycling in the sea. Studies during the past few years have shown that distinct phylogenetic groups of marine heterotrophic bacteria may be responsible for degrading specific types of organic compounds, or for producing specific types of hydrolytic enzymes.
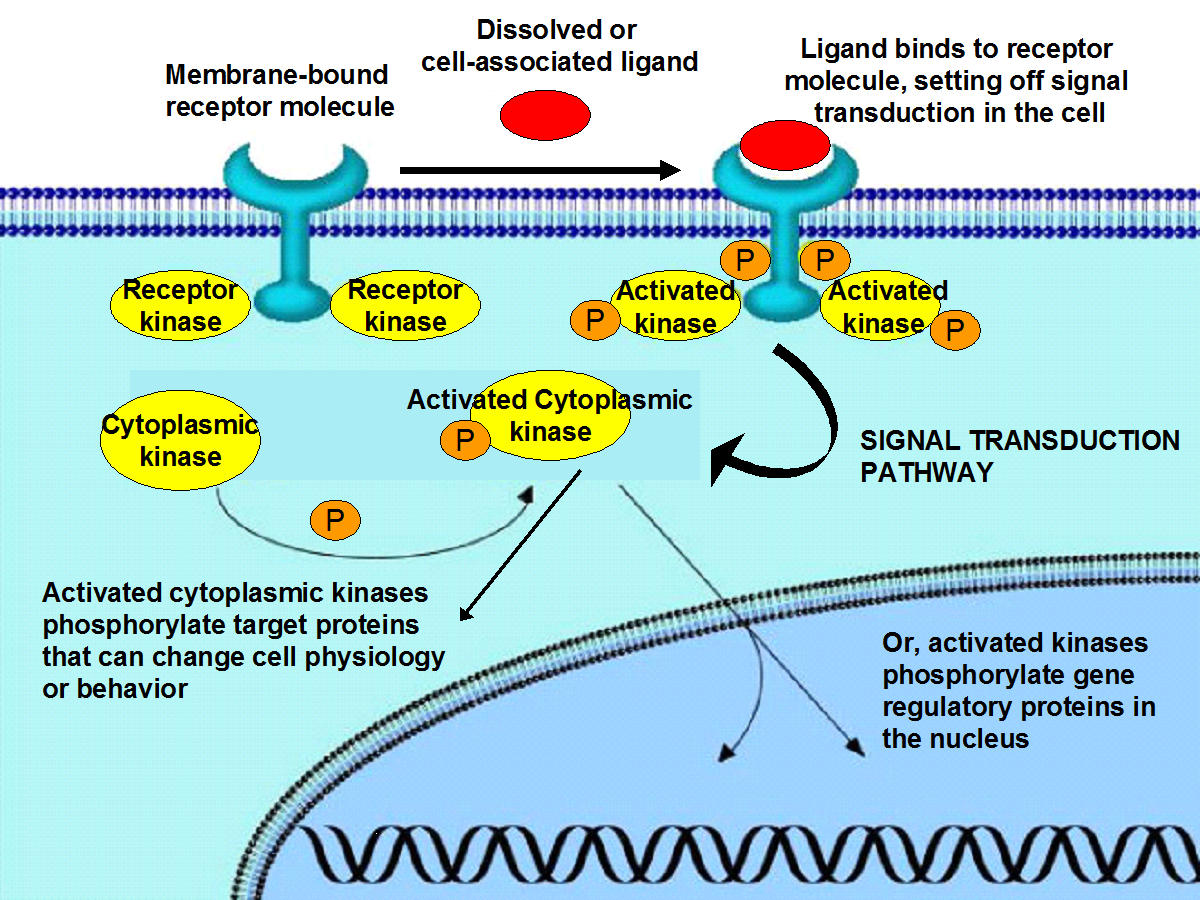
Elucidating the mechanisms by which marine phagotrophic protists might selectively target biochemically, and/or phylogenetically, distinct groups of bacterial prey is thus important for understanding how food web interactions might affect the rate of decomposition of specific types of organic compounds in the sea. Prey selection or rejection is likely to involve universal biochemical pathways: i.e. binding of ligand compounds, for example sugars or peptides, to receptor molecules associated with cell membranes, which triggers intracellular signal transduction pathways leading to changes in cell biochemistry and behavior. We propose an investigation of the role of binding of signal compounds to receptor molecules in protistan cell membranes and of the consequent signal transduction cascades in controlling protistan feeding processes. There is at present very little information on how specific ligand-receptor binding patterns and signal transduction pathways might control the feeding behavior of free-living flagellates, apart from a few studies on laboratory cultured protists. The biochemical mechanisms underlying protistan feeding response is likely to be fundamental to understanding this major top-down control of microbial populations in marine systems. Our proposed study adapts standard protocols of cell biology in a way that could result in new discoveries relevant to marine microbial ecology, and new approaches to assessing the in situ impact of protistan grazing.
This project involved experiments with cultured marine protists. In the first phase of the study, we investigated the potential roles of cell signaling mechanisms in chemosensory response to prey, and in capture of prey cells, by a marine ciliate (Uronema sp.) and a heterotrophic dinoflagellate (Oxyrrhis marina). Inhibition of protein kinase signal transduction biomolecules caused a decrease in both chemosensory response and predation. Inhibition of G-protein coupled receptor signaling pathways significantly decreased chemosensory response but had no effect on prey ingestion. Inhibitor compounds did not appear to affect general cell health, but had a argeted effect. These results support the idea that cell signaling pathways known in other eukaryotic organisms are involved in feeding behavior of free-living protists.
The second phase of this study involved the potential for photosensory behavior in marine protists to aid in detection of algal prey. Expressed rhodopsins were found by proteomic analysis in an investigation of signal receptor molecules in the cell membranes of the marine heterotrophic dinoflagellate Oxyrrhis marina (CCMP604). Protein analyses were done to verify that the rhodopsin molecules expressed by O. marina were in fact present on the outer cell membrane. The initial protein extraction protocol used to evaluate what cell-surface proteins were expressed by O. marina, the Mem-PER protein extraction kit, targeted all membrane proteins (intracellular and plasma membrane proteins). In order to specifically identify those proteins associated with the cell membrane, additional protein extractions were done using Hook Cell Surface Protein Isolation kit (G-Biosciences, Maryland Heights, MO). This protocol uses a biotin-tag/streptavidin affinity column to collect only those proteins exposed on the outer plasma membrane.; Proteins were separated by SDS-PAGE and individual protein bands were excised and digested with trypsin using the In-Gel Tryptic Digestion Kit; Pierce, Thermo Fisher Scientific). Prepared protein samples were anaylyzed by LC-MS/MS at the OSU Mass Spectrometry Facility. Protein data was searched against databases of known proteins using Mascot software (Matrix Science, Boston, MA.)
We inferred these membrane-bound proteins to be sensory rhodopsins, a type of G-protein-coupled receptor trans-membrane signaling molecule. Because phototactic behavior based on sensory rhodopsins has been reported in other protists, we examined the photosensory response of O. marina. We carried out a series of experiments designed to evaluate how O. marina responded to specific wavelengths and intensities of light (movement toward or away from the light source) and to determine whether O. marina could respond to the red fluorescence emitted by blue light excitation of chlorophyll in algal prey. In addition, we tested inhibitors of rhodopsin signal transduction to confirm that the membrane-bound pigments do have a role in behavioral response of the dinoflagellate. To check whether the observed photosensory behavior of O. marina was due to rhodopsin photoreceptors, we used two chemical inhibitors; hydroxylamine and norflurazon. Hydroxylamine has been shown to separate retinylidene chromophores from rhodopsin and inhibit phototaxis in the alga Chlamydomonas reinhardtii and phototaxis in the fungal zoospores of Allomyces reticulates. Norflurazon inhibits the synthesis of carotenoid pigments, such as ß-carotene, used in synthesis of retinal and has been shown to have an effect on microbial phototaxis. As a control, we also tested whether hydroxylamine had an effect on general cell health, O. marina was incubated with hydroxylamine (0.0, 0.25, 0.5, 0.75, 1, and 2 mM) and cell numbers and prey uptake by O. marina were recorded. To determine the effect of hydroxylamanine on O. marina phototaxis, O. marina was incubated with hydroxylamine (0.0, 0.1, 0.12, 0.5, and 1 mM) for 30 minutes in bright light followed by 90 minutes in the dark prior to the start of the phototaxis experiments. The phototaxis index of cells incubated with hydroxylamine was compared to control cells with no hydroxylamine exposure. An additional experiment was done to see if adding all-trans retinal (Sigma-Aldrich) to hydroxylamine treated O. marina could restore phototaxis to levels found in control cells (i.e. not treated with hydroxylamine).
The dinoflagellate exhibited strongest positive phototaxis at low levels (2-3 µE/m(2)/s) of white light when the cells were previously light adapted and well fed. Positive phototaxis was also found for blue (450 nm), green (525 nm), and red (680 nm) wavelengths. In a further test, O. marina showed significantly greater phototaxis toward concentrated algal food illuminated by blue light (to stimulate red chlorophyll-a autofluorescence in the prey), compared with using bleached algae as prey. Concentration of a cytoplasmic downstream messenger molecule, cyclic adenosine monophosphate, a component of the signaling pathway of G-protein-coupled receptor molecules, rapidly increased in O. marina cells after exposure to white light. In addition, treatment with hydroxylamine, a rhodopsin signaling inhibitor, significantly decreased their phototactic response. Our results demonstrate that a heterotrophic marine dinoflagellate can orient to light based on rhodopsins present in the outer cell membrane and may be able to use photosensory response to detect algal prey based on chlorophyll autofluorescence.
In the last year of this project, a post-doctoral colleague, Diego Figueroa, worked in the Sherr Lab (College of Earth, Ocean, and Atmospheric Science, Oregon State University, Corvallis, OR) to survey cultured species of heterotrophic and autotrophic dinoflagellates for the presence of rhodopsin genes.
Type-1 rhodopsin sequences were obtained from Genbank for O. marina, P. lunula, and several protobacteria. These sequences were aligned using ClustalW 1.4 and visually inspected for optimality. Three highly conserved regions within the dinoflagellate sequences that maximized differences with the protobacterial sequences were selected, two near the 3' end of this gene and one near the 5' end. Extraction of DNA was done with Qiagen';s DNAeasy kit. from 15 species of cultured dinoflagellates, including both heterotrophic and autotrophic species: Oxyrrhis marina, Akashiwo sanguinea, Amphidinium longum, Fragilidinium subglobosum, Gymnodinium sp., Gyrodinium sp., Gyrodinium dominans, Glenodinium sp., Lessardia elongata, Peridinium foliaceum, Prorocentrum minimum, Prorocentrum micans, Pyrocystis lunula, and Scrippsiella sp.
Forward primers RHO50 and RHO98 were used each with reverse primer RRHO791 in a Polymerase chain reaction (PCR) for each dinoflagellate culture. PCR product was loaded on a gel for electrophoresis; the gel was then checked under UV light for contamination and consistency of product. The PCR product was cleaned using the standard Montage PCR product cleaning kit. Both the forward and reverse strands were sequenced at the Center for Genome Research and Biocomputing at Oregon State University. The chromatogram for each sequence was visually inspected and the forward and reverse sequences for each specimen were used to generate a consensus sequence. Sequences were aligned using ClustalW 1.4 and visually inspected for optimality. The sequences were then translated into their corresponding amino acid structure.
A 730 bp sequence was obtained for 8 of the dinoflagellates (O. marina, P. foliacium, L. elongata, G. dominans, Gyrodinium sp., Gymnodinium sp., Scroppsiella sp., and P. lunula) with primer pair RHO50 and RHO791. This primer pair was not effective with the other 6 dinoflagellates. A 650 bp sequence was obtained from A. sanguinea using primer pair RHO98 and RHO791. For the remaining 5 dinoflagellates (A. carterae, F. subglobosum, Glenodinium sp., P. minimum and P. micans) the PCR product was fragmented when using either pair of primers. The fragments were observed under UV light after gel-electorphoresis; they ranged from around 10bp to 300bp in length. Using Invitrogen's gel extraction/PCR purification kit, we were able to sequence the larger fragments, recovering a 260 bp sequence for all 5 dinoflagellates. All the sequences showed high similarity, being almost identical throughout the first 400bp. The recovered sequences were more variable at the 3' end.
This ubiquitous dinoflagellate rhodopsin gene was probably acquired by the ancestor to all dinoflagellates from protobacteria through lateral gene transfer. A similar pattern is found in the gene coding for the photosynthetic enzyme Rubisco of autotrophic dinoflagellate which is of prokaryotic (form II) instead of eukaryotic (form I). A recent Nature Communications publication (Slamovitz et al. 2011) proposed that the major rhodopsin in dinoflagellates is likely used as a proton pump to enhance acidity in food vacuoles, although sensory rhodopsins are also present in dinoflagellate genomes. If so, then our results are relevant to the more common proton pump rhodopsin.
A manuscript based on this work is in preparation (Figueroa, et al. 2012). Sequences for the dinoflagellate rhodopsin genes will be submitted to GenBank and the accession numbers recorded at BCO-DMO when that paper is submitted.
Related Documents:
Sherr E.B. and B.F. Sherr. 2007. Heterotrophic dinoflagellates: a significant component of microzooplankton biomass and major grazers of diatoms in the sea. Mar. Ecol. Prog . Ser. 352: 187-197
Sherr E.B. and B.F. Sherr. 2009. Capacity of herbivorous protists to control initiation and development of mass phytoplankton blooms. Aquatic Microbial Ecology 57: 253-262.
Publications:
Hartz, A.J., B.F. Sherr and E.B. Sherr. 2008 Using Inhibitors to investigate the involvement of cell signaling in predation by marine phagotrophic protists. J. Euk. Microbiol. 55:18-21
Hartz, A.J., B.F. Sherr and E.B. Sherr. 2011. Photoresponse in the heterotrophic marine dinoflagellate Oxyrrhis marina. J. Euk. Microbiol.58:171-7.
Figueroa, D., B.F. Sherr and E.B. Sherr. (in preparation, 2012) Proteobacterial rhodopsin gene is ubiquitous in marine dinoflagellates.
| Dataset | Latest Version Date | Current State |
|---|---|---|
| Links to GenBank accession numbers for 18S rDNA from the ciliate, Euplotes nobilii, sequenced by the Sherr lab at OSU, Corvalis, OR in 2012 | 2012-04-13 | Final with updates expected |
Principal Investigator: Barry Sherr
University of Oregon
Co-Principal Investigator: Evelyn Sherr
University of Oregon